This information was most recently updated November 9, 2006.
Although all cells possess a large number of different types of repair system, each relatively specific for a certain type of DNA damage, those repair systems can be grouped into four broad categories:
Although it might seem that direct reversal of damage would be the simplest way to correct the damage, in most cases the reverse reaction is not possible for thermodynamic or kinetic reasons. In a few cases, the reaction is reversible, and in some of these cases mechanisms have been developed to take advantage of that reversibility.
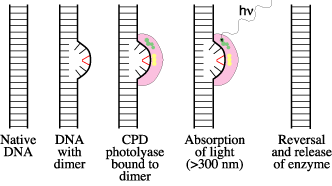
The best studied of these is photoreversal of CPDs by the enzyme, CPD photolyase. The reaction catalyzed by CPD photolyase, called "photoreactivation", was the first DNA repair process to be discovered. It was discovered by bacteriophage geneticists in 1949, well before the double-helical structure of DNA was discovered in 1953.
CPD photolyase contains two chromophores responsible for absorbing light energy. In all such photolyases, one of the chromophores is FADH-, and the other is either methenyl-tetrahydrofolate (MTHF) or 8-hydroxy-5-deazaflavin (8-HDF). MTHF and 8-HDF act as primary light gatherers (colored green in the diagram below), transferring their energy to FADH- (colored yellow in the diagram). The energy from FADH- is then used to split the dimer:
CPD photolyases are found in bacteria, fungi, plants and many vertebrates, but not in placental mammals. In addition, 6-4 photolyases (which repair 6-4PPs) have been found in insects, reptiles and amphibians, but not in E. coli, yeast or mammals.
It is interesting that the sensitivity of mouse skin to UV damage could be largely corrected by genetically engineering the mice to express a foreign CPD photolyase gene. A foreign 6-4 photolyase gene was much less effective. These results suggest that production of CPDs is a more serious problem for mice than production of 6-4 photoproducts.
Another example of direct damage reversal is repair of O6-alkyl guanine by transfer of the alkyl group from the DNA to a cysteine in a protein, an O6-alkylguanine-DNA alkyltransferase. The alkyl groups handled by this protein range in size from methyl to benzyl. Homologues of this protein are present in all living organisms. Unfortunately, once the alkyl group has been transferred to the protein, the protein is permanently inactivated (alkyl cysteine is very stable). Additional repair requires the synthesis of new protein molecules. To help accomplish this, in E. coli and perhaps in other organisms, the alkylated version of the protein induces increased transcription of the gene encoding the protein. Epigenetic silencing of the human gene encoding this protein leads to elevated mutation frequency and increased susceptibility to cancer.
A third example of direct damage reversal is provided by AlkB, a protein that is found in most, perhaps all, living organisms (including humans). This protein is a member of the class of enzymes called alpha-keto-glutarate-dependent and iron-dependent oxygenases (aKG-Fe(II)-oxygenases), which use iron-oxo intermediates to oxidize chemically inert compounds. In the process, alpha-keto-glutarate is converted to succinate and CO2, which is similar to one of the steps in the tricarboxylic acid cycle. AlkB is capable of reversing methylation at the 1 position of adenine and the structurally similar 3 position of cytosine, as shown in this diagram:
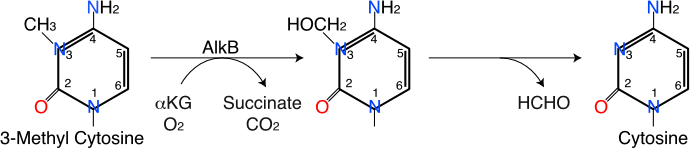
The reaction catalyzed by AlkB couples the oxidative decarboxylation of alpha-keto-glutaric acid to the hydroxylation of the methyl group. The methyl group then spontaneously decomposes to formaldehyde, restoring the original base.
A final example of direct damage reversal is the sealing of a subset of nicks in DNA by DNA ligases. Of course, DNA ligases can only seal nicks having 5'-phosphates and 3'-hydroxyls. Nicks with other configurations, or nicks accompanied by additional backbone or base damage, require more complicated processing prior to repair and would not be classified as direct damage reversal mechanisms.